我們在2014年的12月11日,邀請了中研院農生中心的常怡雍老師來跟大家分享他在葉綠素代謝上的新發現。
常老師在找尋植物的耐熱基因上卓然有成,這次卻不講他的耐熱基因們,固然是因為謙虛(常老師說,既然筆者的名字跟葉綠素很像,他又剛好有這個新發現,所以他覺得一定要講XD),但其實這個新發現還是從耐熱基因的研究產生的。
怎麼說呢?原來,一開始這個突變株是被命名為dlt4(defect in long-term acquired thermotolerance)。研究團隊多年來一直都是以阿拉伯芥(Arabidopsis thaliana)為材料,找尋暴露在高溫下之後,重回正常狀態培養發生漂白化(bleaching)的突變株,而dlt4就是其中之一。
研究團隊找到有合適的性狀的突變株之後,接著就是去找突變的基因。當他們找到dlt4發生突變的位點以後,卻發現那個基因的序列跟藍綠菌的chlorophyll synthase(葉綠素合成酶)很像(以下改稱dlt4為chlg)。
研究團隊接著發現,正如上圖(b)所顯示的,chlg-1突變株的漂白化性狀,如果在熱處理後回到黑暗中培養兩天,就不會出現了;可是他們在阿拉伯芥突變株種源庫中找到的相同基因的突變種(稱為chlg-2),卻是不管有沒有熱處理都會有漂白化的性狀。
原來,chlg-1(原來的dlt4)突變株的葉綠素合成酶上發生的突變是所謂的「錯義突變」(missense mutation)。這個突變使得原來葉綠素合成酶的第217號氨基酸由甘氨酸(glycine,縮寫為G)變為精氨酸(arginine,縮寫為R)。由於甘氨酸與精氨酸的性質非常不同,大小差距也非常大(如下圖),造成整個葉綠素合成酶變得不穩定,結果就是chlg-1突變株的葉綠素合成酶的量只有野生種的十分之一而已。
而葉綠素合成酶的量下降到原來的一成,使得chlg-1突變株在熱處理後發生植醇葉綠素 a(chlorophyllide a)累積的現象。不過,在植物裡面,有兩種狀況下會產生植醇葉綠素a。
哪兩種狀況呢?一種是從新(de novo)合成葉綠素a的途徑,這部分已知是由原植醇葉綠素氧化還原酶(protochlorophyllide oxidoreductase)負責;另一種則是在第二光系統(photosystem II)中的D1蛋白分解的時候。由於chlg-1的白化苗在熱處理後再照光,並不會出現漂白化現象,顯示問題應該不是出在從新(de novo)合成葉綠素a。為什麼呢?原來,從新合成葉綠素a需要光來啟動(沒有光,就不需要合成葉綠素a對吧?);所以,既然突變株的白化苗在熱處理之前不照光就不會漂白化,表示跟從新合成無關。
而植醇葉綠素a的累積,為什麼會導致漂白化呢?原來植醇葉綠素a的累積,會產生自由基,對植物造成傷害。而第二光系統的D1蛋白質每30分鐘就要更換一次,每次的更換,整個第二光系統要先拆解,除了更換D1蛋白以外,裡面的葉綠素a也會被分解為植醇葉綠素a與植醇,然後再重新組合...目前認為CHLG蛋白應該跟這部分有關係,所以幼苗熱處理之後,如果放在黑暗中兩天,因為沒有光,也不能進行光合作用;雖然已經有第二光系統的存在,但是沒有光也不用更換D1蛋白,所以就不會產生植醇葉綠素a,也不會有造成傷害的自由基囉!
所以,常老師的研究團隊原本是在找尋跟耐高溫逆境有關的突變株,最後卻找到了缺乏它使植物遇到高溫會白化,但跟耐高溫逆境無關的基因;怎麼會這樣呢?
事實上,光是植物,除了常老師研究團隊的這個例子以外,筆者就可以想到大名鼎鼎的det2,當初是為了要找在「去白化」(de-etiolation)這件事上面有障礙的基因,結果det2卻是合成芸苔素(brassinolide)的酵素;還有多年前好幾個不同實驗室想要找到可以逆轉光敏素B突變株的性狀,卻找到了許多與生長素(auxin)合成有關的基因...這只能說,生物原本就很複雜,當我們以「性狀」(phenotype)來做為篩選的基準時,就好比我們要一群人從花蓮出發到台北,最後到不了台北的人,不見得都是買不到北迴線車票的人一樣...雖然北迴線真的很難很難很難買,哈!
參考文獻:
Yao-Pin Lin, Tsung-yuan Lee, Ayumi Tanaka, Yee-yung Charng. 2014. Analysis of an Arabidopsis heat-sensitive mutant reveals that chlorophyll synthase is involved in reutilization of chlorophyllide during chlorophyll turnover. The Plant Journal. 80(1):14–26.
常老師在找尋植物的耐熱基因上卓然有成,這次卻不講他的耐熱基因們,固然是因為謙虛(常老師說,既然筆者的名字跟葉綠素很像,他又剛好有這個新發現,所以他覺得一定要講XD),但其實這個新發現還是從耐熱基因的研究產生的。
怎麼說呢?原來,一開始這個突變株是被命名為dlt4(defect in long-term acquired thermotolerance)。研究團隊多年來一直都是以阿拉伯芥(Arabidopsis thaliana)為材料,找尋暴露在高溫下之後,重回正常狀態培養發生漂白化(bleaching)的突變株,而dlt4就是其中之一。
 |
| (a)高溫處理過的chlg-1(dlt4-1),出現漂白化現象。 (b)如chlg-1在高溫處理過後,回到黑暗中培養二天,則無漂白化現象發生。 圖片來源:The Plant Journal |
研究團隊接著發現,正如上圖(b)所顯示的,chlg-1突變株的漂白化性狀,如果在熱處理後回到黑暗中培養兩天,就不會出現了;可是他們在阿拉伯芥突變株種源庫中找到的相同基因的突變種(稱為chlg-2),卻是不管有沒有熱處理都會有漂白化的性狀。
原來,chlg-1(原來的dlt4)突變株的葉綠素合成酶上發生的突變是所謂的「錯義突變」(missense mutation)。這個突變使得原來葉綠素合成酶的第217號氨基酸由甘氨酸(glycine,縮寫為G)變為精氨酸(arginine,縮寫為R)。由於甘氨酸與精氨酸的性質非常不同,大小差距也非常大(如下圖),造成整個葉綠素合成酶變得不穩定,結果就是chlg-1突變株的葉綠素合成酶的量只有野生種的十分之一而已。
 |
| 甘氨酸。圖片來源:wiki |
{kind=link}
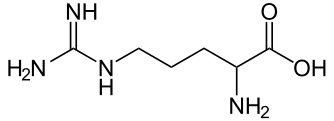 |
| 精氨酸。圖片來源:wiki |
{kind=link}
 |
| 葉綠素a(chlorophyll a)。 上面的多角形環狀結構就是植醇葉綠素a, 下面長長的碳鏈是植醇(phytol)。 圖片來源:wiki |
{kind=link}
而植醇葉綠素a的累積,為什麼會導致漂白化呢?原來植醇葉綠素a的累積,會產生自由基,對植物造成傷害。而第二光系統的D1蛋白質每30分鐘就要更換一次,每次的更換,整個第二光系統要先拆解,除了更換D1蛋白以外,裡面的葉綠素a也會被分解為植醇葉綠素a與植醇,然後再重新組合...目前認為CHLG蛋白應該跟這部分有關係,所以幼苗熱處理之後,如果放在黑暗中兩天,因為沒有光,也不能進行光合作用;雖然已經有第二光系統的存在,但是沒有光也不用更換D1蛋白,所以就不會產生植醇葉綠素a,也不會有造成傷害的自由基囉!
 |
| 藍綠菌的第二光系統,D1蛋白以深藍色標出, 葉綠素a為黃綠色標示。 圖片來源:wiki |
{kind=link}
所以,常老師的研究團隊原本是在找尋跟耐高溫逆境有關的突變株,最後卻找到了缺乏它使植物遇到高溫會白化,但跟耐高溫逆境無關的基因;怎麼會這樣呢?
事實上,光是植物,除了常老師研究團隊的這個例子以外,筆者就可以想到大名鼎鼎的det2,當初是為了要找在「去白化」(de-etiolation)這件事上面有障礙的基因,結果det2卻是合成芸苔素(brassinolide)的酵素;還有多年前好幾個不同實驗室想要找到可以逆轉光敏素B突變株的性狀,卻找到了許多與生長素(auxin)合成有關的基因...這只能說,生物原本就很複雜,當我們以「性狀」(phenotype)來做為篩選的基準時,就好比我們要一群人從花蓮出發到台北,最後到不了台北的人,不見得都是買不到北迴線車票的人一樣...雖然北迴線真的很難很難很難買,哈!
參考文獻:
Yao-Pin Lin, Tsung-yuan Lee, Ayumi Tanaka, Yee-yung Charng. 2014. Analysis of an Arabidopsis heat-sensitive mutant reveals that chlorophyll synthase is involved in reutilization of chlorophyllide during chlorophyll turnover. The Plant Journal. 80(1):14–26.
留言
張貼留言